
Networked Pressures
plant breeding from the process-oriented, topsy-turvy worldview of ecology

“…the topsy-turvy world of ecology appears to stand all the fundamental assumptions of normal science on their heads. Instead of a world that is closed, atomistic, reversible, deterministic and universal; we now see one that is (respectively) open, organic, historical, contingent and granular.” - Dr. Robert Ulanowicz
One thing plant breeders do is apply selection pressure. Selection pressure is thought of as the external forces acting on an organism that affect its survival, such as predation, resource competition, pathogen exposure, heat or cold, and so on. Human-mediated selection pressure, Darwin referred to as artificial selection. Judgements like “this plant is too spiny, so I’ll cut it down” or “this fruit is sweet, so I’ll spread the seeds”, and so on. Joseph Lofthouse, author of the excellent book Landrace Gardening, feels that 85% of selection is done by the ecosystem. The other 15% is the space for directed breeding work aka artificial selection. What would plant breeding look like from the process-oriented, “topsy-turvy” view of ecology ?
The systems ecologist Dr. Robert Ulanowicz uses the ecological concept of autocatalysis as didactic entry point into this view. A simple depiction of autocatalysis is the interaction of the elements A, B, and C in a loop. Any change causing an increase in one element ripples through the network and back on itself, causing more and more resources to be pulled into the system. This reveals a level of selection pressure acting top-down through network feedback dynamics. These networks are very common in ecology.
“Autocatalysis is capable of exerting selection pressure upon its ever-changing, malleable constituents.” - Dr. Robert Ulanowicz (source)
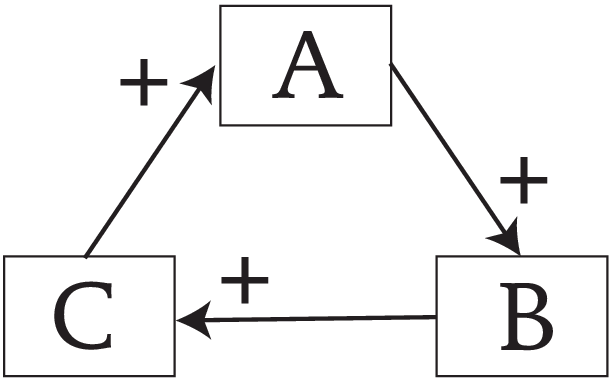
We are taught not to think using circular logic. Deductive reasoning is the gold standard, while circularity synonymous with nonsense. Yet if we abandon circular causation completely, we only see the mechanical subsets of living systems, overlooking the non-mechanical behaviors they exhibit. In terms of selection pressure, the conventional emphasis is on linear, individual-based, and competition-driven processes. The ecological perspective integrates this with broader non linear, network-based, cooperative dynamics.
Plant breeding influenced by ecological thinking places a greater focus on growth and development at multiple scales. This can apply to the level of the genome, the landscape and the ecosystem. It can mean breeding for polycultures, agroforestry systems, and ecosystem services. It can involve adaptive management, community engagement, and the holistic evaluation of traits. It might even mean selecting for emergent properties that we hadn’t foreseen.
Let’s take a look at how living systems grow and develop through autocatalysis (source). The representation below compares a young system A to an older version of the system B.
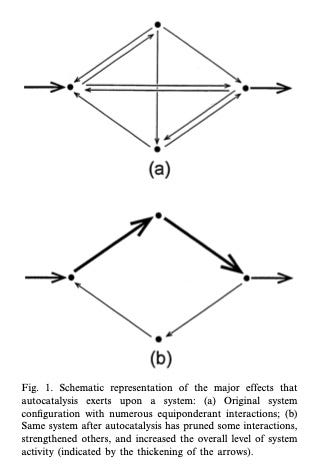
We notice right away that there are many more connections in the original system A compared to the more streamlined configuration of B. Many connections are pruned away in the development from A to B, yet the total amount of material and energy throughput is increased, as indicated by the thicker arrows. At the level of the brain, it represents the changes from a child to an adult. For a plant, it’s a young seedling compared to an older flowering plant. At the level of the ecosystem, this represents what we call succession. As ecosystems climb this developmental curve, there is a sweet spot they tend to hover around, having to do with the trade-off between resiliency (or diversity) and overall productivity (or efficiency). From time to time, the system gets challenged by what ecologists call perturbations. External shocks occasionally disrupt the system, causing it to reset to an earlier developmental phase. The stronger the stress, the further the reset goes back in developmental time, resembling to a greater degree system A. In a terrestrial ecosystem, this results in the proliferation of early succession plants that we colloquially call weeds. A stronger perturbation can set succession back even further to lichens. Whereas an old growth forest is networked like system B—productive, robust, with relatively stable and determinant flows.
When we prune a plant, it sends out new green shoots. Before sending out new shoots, it forms a layer to heal the wound called callus tissue. The callus tissue is sort of like an undifferentiated blob of gel. As the counterpart to stem cells in animals, callus has the potential to develop into any kind of plant tissue. Callus tissue is also formed when plants heal graft unions. Callus tissue has very high plasticity, just like system A in the schematic above. In fact, sometimes simply pruning a shoot can cause a doubling of the genome, or polyploidy (source). Variation of chromosome numbers is found naturally in callus tissue, an indicator of its high plasticity. A plant that is flowering is on the other end of the spectrum like system B.
Callus tissue also forms when growing plants in tissue culture. The cytogeneticist Barbara McClintock received a Nobel prize in 1983 for the discovery of transposons. Transposons are the cellular mediators of plasticity (system A), and activate after stress. The title of McClintock’s Nobel speech is called The Significance of Responses of the Genome to Challenge. Various synonyms are used for challenges to a system: stress, disturbance, perturbation. Another term that McClintock used is shock, known as her “genomic shock hypothesis.” In the Nobel speech, McClintock talks about the novel mutations that arise from growing out tissue cultured corn plants. The title of the speech hints that there’s a deeper significance of the responses to genome shock than we had realized. The significance has continued to reveal itself as scientists have gradually ditched the term “junk DNA” to refer to transposons. There has been an increasing realization is that transposons play an important role in genome regulation and contribute to evolutionary processes. When an organism is more developed (as in system B), transposons are inactive, kept silent by epigenetic mechanisms. But when a challenge arises for the organism, the transposons mobilize. They jump around the genome, cut- and copy-pasting themselves, hence the name “jumping genes.” They have also been called the “weeds of the genome.” By the ecological definition of a weed, it’s an accurate metaphor in that they come out of hiding and proliferate after disturbances. Weeds can also direct the system into different future states, as transposons do with the genome. Weeds sometimes spread to new ecosystems, through close contact or over larger distances via an intermediary such as a bird. Likewise, transposons can spread into new genomes, through close contact or via an intermediary such as a virus.
The notion of graft hybridization first appears in old Chinese texts. There are many examples of grafting distantly related plants for their combined qualities. For example, there is the recommendation to graft pear onto jujube to produce the best flavored pears. In the West there has traditionally been much skepticism toward graft hybrids. One reason for this is that some researchers early on didn’t apply the proper methods to maximize the chances of success. Under the normal technique of graft propagation these effects go unnoticed to a large extent. In 1907, German botanist Hans Winkler demonstrated that you can create graft chimeras by grafting then making a cut at the graft union. Occasionally a chimeric shoot arises from the graft union. It consists of a combination of two different types of cells growing together, which in their most harmonious pattern occupy adjacent layers like a hand in glove (a periclinal chimera). They form a unit, but are able to be separated again. After Winkler demonstrated the nature of graft chimeras, for many people this settled the question of graft hybrids. They decided graft chimeras must be the sole phenomenon responsible for the graft hybrid debate, and indeed this is still a prevalent view today among horticulturists and plant breeder (source). There is some irony to this, as Winkler himself was not convinced this was the case. Winkler saw clearly that there were 3 theoretically possible types of graft hybrids: 1) fusion hybrids 2) influenced hybrids and 3) chimeras. In one of his experiments synthesizing graft chimeras, Winkler found a polyploid shoot coming out of a graft union that did not appear to be a chimera, but a cell fusion hybrid between the two species. This plant unfortunately died before it could be propagated out to other researchers, and with the same wave of the hand, many commentators brushed this off also as a probable chimera (source). Nearly a century later in 2014, cell fusion hybrids were selected out from the graft union of two different Nicotiana species, see Horizontal genome transfer as an asexual path to the formation of new species. Winkler’s cell fusion hybrids at the graft union have now been confirmed and created experimentally along with graft chimeras.
So what about the “influenced” graft hybrids? Winkler only noted the theoretical possibility of these, but his experimental method could not produce them, so he remained agnostic about their existence. Unlike chimeras or cell fusion hybrids, influenced graft hybrids do not arise from a shoot at the graft union. Ivan Michurin developed a technique to produce influenced graft hybrids called the mentor graft. Here is a depiction, from the study “Hereditary changes in Capsicum annuum LI Induced by ordinary grafting.”
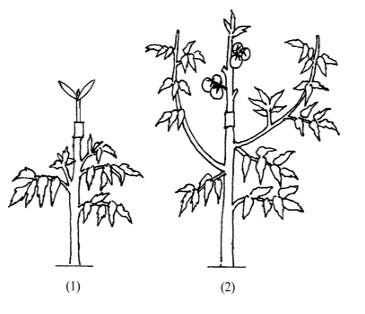
The technique is to graft a young hybrid seedling onto older rootstock, then while it grows prune all but the 2-3 leaves at the scion’s tip. In this way the scion (and its seedlings) will be “influenced” (or mentored as Michurin called it), with an increasing intensity over time, by a mostly one-directional flow from the rootstock. Like the other types graft hybrids, influenced graft hybrids have likewise been confirmed in the literature (e.g. Molecular mechanism of graft transformation in Capsicum annuum), but unlike the other two types these still elicit incredulity. In fact, there are more than two dozen studies confirming these graft hybrids (including a few showing genetic influence across the layers of chimeras). These studies frequently use Michurin’s mentor grafting technique but not always. There is some variation in the experimental designs as well as kinds/intensity of effects. These experiments are typically set up so that the trait transfer is obvious to notice, such as a green leaf scion growing on a purple leaf rootstock, or a jalapeño pepper on a bell pepper rootstock*.
Mentor grafting is about affecting an increased flow from an older, larger, stabilized plant (system B) into a new, small, plastic seedling (system A). Stressful conditions combine in the young organism, which mobilize transposons. Spreading around the genome they disrupt genes, create new regulatory networks, and introduce new traits by causing mutations, gene duplications, and chromosome rearrangements. Additionally, in the context of a mentor graft, transposons can promote horizontal genetic transfer through the genome instability and repair mechanisms they trigger. You could reason autocatalytic loops at play in this situation, for example foreign genetic material (such as RNAs) activate transposons in the scion, the resulting genomic instability promotes the uptake of foreign genetic material, which then continues to activate transposons. The leaf pruning stress of the scion further engages types of these loops, for example by increasing extracellular vesicle production, transport and uptake. This is like a chaos theory approach to plant breeding: a new system with unstable initial conditions is fitted within the highly perturbative flows of a large attractor. The intensity and the type of these conditions can be varied to affect the results. In some experimental conditions, the variation does not show up until seed of the scion are grown.
Applying the ecological view to plant breeding and development can be taken in two directions. One direction is taking plant selection out into multiple layers of the ecology and the community to benefit both. The other direction is toward an ecology of the genome, applying disturbance regimes and nurturing new developmental pathways for organism similar to the management of the wider ecosystem. Working with disturbance regimes and directing development on both of these levels holds great potential for synergistic interactions. Returning to the ideas of selection pressure and autocatalysis, they can also form a loop. In the context of a mentor graft, diverse foreign biomolecules flood into the scion in a sustained way. These stress conditions are a form of selection pressure, one which not only acts to select among the existing diversity, but also reconfigures the existing diversity, and is source of new diversity. This is essentially applying autocatalysis to selection pressure. As chain reactions take place, the more dynamic the results can be. This dynamic form of selection pressure could be referred to as evolutionary catalysis, capturing the ideas of rapid trait development and an increased genetic pool.
Regarding the kinds of molecules that move across the graft union, there are many: “macromolecules such as proteins and hormones, as well as small signaling molecules such as sRNAs and miRNAs, can be exchanged. Further, cpDNA undergoes genetic material exchange, and horizontal transfer of nuclear genes occurs at the graft union” (source). What’s more, even molecules that have been considered too large to move across the graft union such as chromatin masses have been shown to do so, enhanced by the technique of pruning the scion leaves (source). Chromatin is a relatively large and complex macromolecule consisting of DNA, RNA, and protein.
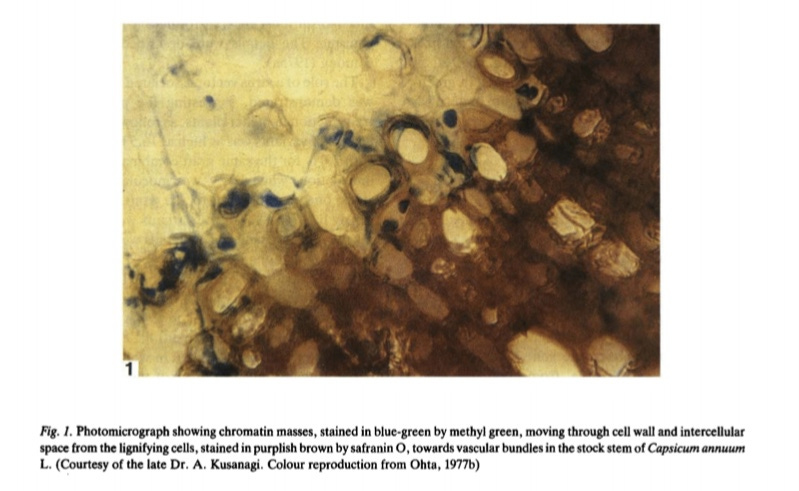

The ability of chromatin to travel suggests that other large biomolecules, such as proteins, large RNAs, and even organelles, might also translocate under similar stress conditions. Stress conditions are able to increase the permeability of the graft union and dilate the pathways that these molecules move thorough. These conditions promote the horizontal transfer of epigenetic modifications, genetic material, microbes, and other information. We might characterize a graft hybrid as a horizontally integrative organism.
The photo below shows a graft hybrid of a grapevine grown on the very distantly related Shisandra rootstock (source). G is the original unmodified green grape cultivar, while H is the same cultivar after growing and transforming for several years on Shisandra rootstock. The grapes became much darker and closer in color to shisandra berries, and they also now contain the medicinal components from shisandra. All of the traits stabilized, even when the Vs (Vitis on Shisandra) was grown out later on its own roots.
Stay tuned for more thoughts on symbiogenic and process-oriented approaches to plant breeding.


*some of the studies done on influenced graft hybrids:
Strou, M. et al. 1963. Modifications transmitted to the offspring, provoked by heterograft in Solanum melongena.
Tsaballa, A. et al. (2013). Molecular studies of inheritable induced changes in pepper (Capsicum annuum) fruit shape.
Zhang, S. et al. (2024). Interfamily Grafted Hybrids Vitis vinifera/Schisandra chinensis Resulted in Transcriptomic, Phenotypic, and Metabolic Changes
Li, J. et al. (2013). Heritable variation and small RNAs in the progeny of chimeras of Brassica juncae and Brassica oleraceae.
Hirata, Y. et. al. (1994). Genetic constitution of germ cells in intervarietal and interspecific chimeras of Brassica induced by in-vitro grafting
Pan Xiang-wen, et al. (2012). Optimization of Distant Graft Mutagenesis Technology in Soybean
Zhou, X. et al (2013). Confirmation of a purple-leaved plum graft hybrid.
Frankel, R. (1962). Further evidence on graft induced transmission to progeny of cytoplasmic male sterility in Petunia.
Yagishita, N., & Hirata, Y. (1987). Graft-induced change in fruit shape in Capsicum annuum LI genetic analysis by crossing.
Yagashita, N. and Y. Hirata, 1984. Genetic analysis of new cultivar obtained by grafting in Capsicum annuum
Hirata, Y. (1980). Graft-induced changes in eggplant (Solanum melongena L.), 2: Changes of fruit color and fruit shape in the grafted scions and in the progenies from the grafted scions.
Excellent essay, the plant breeding world needs more of this deep thought. Mentor grafting is criminally unknown; I hope this article helps publicise it.
Great essay! I’ve been wanting more information on mentor grafting and here you were. I’m curious to try it on Dioscorea bulbifera on D. polystachya and purple D. alata, jackfruit on fig and water apple on rose apple.